


ЕЭШмбѕЯТЯѕЛЏЩњЮяФЄжаОњШКЕФОКељдіжГФЃаЭ
дкЩњЮяЯѕЛЏЙ§ГЬжаЃЌАБЪзЯШгЩАБбѕЛЏОњбѕЛЏЮЊбЧЯѕЫсбЮЃЌШЛКѓгЩбЧЯѕЫсбѕЛЏОњбѕЛЏЮЊЯѕЫсбЮЃЌећИіЯѕЛЏЗДгІЪЧСНжжОњШКаЭЌзїгУЕФНсЙћЁЃвђДЫЃЌОњШКжЎМфЕФЩњГЄКЭЛљжЪзЊЛЏЦНКтжСЙиживЊЃЌЗёдђНЋдьГЩжаМфВњЮябЧЯѕЫсбЮЕФЛ§РлЁЃСэвЛЗНУцЃЌЭЈЙ§ЖдЯѕЛЏЗДгІЕФСНДѓОњШКНјааКЯРэЕФЕїПиЃЌЪЙЯѕЛЏЙ§ГЬВњЩњГжОУЮШЖЈЕФбЧЯѕЫсбЮЛ§РлВЂжБНгНјааЗДЯѕЛЏЃЌаЮГЩЖЬГЬЯѕЛЏ—ЗДЯѕЛЏЃЌдђПЩвдДяЕНЃл1ЁЂ2ЃнЃКЂйдкКУбѕНзЖЮПЩМѕЩй25%ЕФашбѕСПЃЛЂкдкШБбѕНзЖЮПЩМѕЩй40%ЕФгаЛњЬМдДЃЛЂлбЧЯѕЫсбЮЕФЗДЯѕЛЏЫйТЪЭЈГЃБШЯѕЫсбЮИп1.5ЁЋ2.0БЖЃл3ЃнЁЃетЖдДѓСПЕФИпАБЕЊЕЭЬМдДЗЯЫЎДІРэОпгаЪЎЗжживЊЕФвтвхЁЃЪЕЯжбЧЯѕЫсбЮЛ§РлЕФЗНЗЈгагЮРыАБвжжЦЁЂДПжжХрбјЁЂЮТЖШбЁдёЕШЃЌЕЋЫќУЧЖМжЛЪЪгУгквЛЖЈЕФЗЖЮЇЁЃЭЈЙ§ПижЦЗДгІЬхЯЕжаШмНтбѕХЈЖШПЩдкЩњЮяФЄЗДгІЦїжаЪЕЯжГжОУЮШЖЈЕФбЧЯѕЫсЛЏЃЌЦфдвђЪЧдкЯѕЛЏЩњЮяФЄжаДцдкзХАБбѕЛЏОњКЭбЧЯѕЫсбѕЛЏОњжЎМфЖдШмНтбѕЕФОКељРћгУЃЌгЩДЫдьГЩЩњЮяФЄжаОњШКжЎМфЕФОКељЩњГЄЁЃСНжжОњШКдкЯѕЛЏЩњЮяФЄжаЕФПеМфОКељЬиЕуЃЌЖдЗДгІЦїЕФЩшМЦвдМАЙЄГЬВйзїЖМгаКмживЊЕФвтвхЁЃ
БОЮФФПЕФЪЧдкЕЭШмНтбѕЬѕМўЯТЃЌНЈСЂЩњЮяСїЛЏДВФкЯѕЛЏЩњЮяФЄжаСНжжОњШКЕФОКељдіжГФЃаЭЃЌгУвдНтЪЭЯѕЛЏЗДгІЩњЮяФЄжаАБбѕЛЏОњКЭбЧЯѕЫсбѕЛЏОњжЎМфЕФПеМфЗжВМКЭЖдЛљжЪЕФОКељРћгУЁЃ
ЯжгаЕФЩњЮяФЄФЃаЭДѓЖрЮЊЮШЬЌФЃаЭ[4]ЃЌШЯЮЊЩњЮяФЄЕФаджЪ(ШчЩњЮяФЄЕФКёЖШЁЂЛюад)ЁЂЛљжЪКЭЮЂЩњЮяжжШКдкЩњЮяФЄжаЕФЗжВМЖМЪЧКуЖЈЕФЁЃетаЉФЃаЭМАЫљЕУНсТлМЋДѓЕиЗсИЛСЫЩњЮяФЄРэТлЃЌЕЋЪЕМЪЩЯМДЪЙЗДгІЦїДІдкЮШЬЌЬѕМўЯТЃЌЩњЮяФЄЕФзщГЩЁЂКёЖШМАжжШКЗжВМвВдкВЛЖЯБфЛЏЁЃОнДЫЃЌWannerЕШШЫНЈСЂСЫЗЧЮШЬЌЩњЮяФЄФЃаЭЃл5ЁЋ7ЃнЃЌЫќПЩвддЄВтЩњЮяФЄКёЖШЕФБфЛЏКЭУшЪіЮЂЩњЮяжжШКМАЛљжЪдкЩњЮяФЄжаЕФЗжВМЃЌФПЧАБЛЙуЗКВЩгУЃЌЕЋЪЧWannerФЃаЭгавдЯТМИЗНУцЕФШБЯнЃКЂйWannerФЃаЭЪЧЦНАхФЃаЭЃЌМДШЯЮЊжЇГжНщжЪПЩвдПДзївЛИіЦНАхЃЌЖјдкЩњЮяСїЛЏДВФкВЩгУЮЂСЃзїЮЊжЇГжНщжЪЃЌЦфГпЖШгыЩњЮяФЄЮЊЭЌвЛЪ§СПМЖЃЌетШнвздьГЩКмДѓЕФЮѓВюЃЛЂкWannerФЃаЭШЯЮЊећИіЩњЮяФЄЖМОпгаЛюадЃЌЕЋЪЧЫцзХЮЂаЭДЋИаММЪѕЕФЗЂеЙЃЌжЄУїдкЩњЮяФЄжаO2ЕФРЉЩЂЩюЖШЭЈГЃЮЊ100μmзѓгвЁЃвђДЫЃЌКУбѕЩњЮяФЄжаОпгаЛюадЕФВПЗжвВЮЊЯрЭЌЕФЪ§СПМЖЃЛЂлWannerФЃаЭНсЙЙИДдгЃЌвђЫиЙ§ЖрЃЌМЦЫуЙ§ГЬЗБЫіЃЌашвЊзЈУХЕФШэМўМЦЫуЧѓНтЃЌКмФбЪЕМЪгІгУЁЃ
1БэУцЛюадФЄРЉеХФЃаЭЕФНЈСЂ
1.1ЛљБОЫМТЗ
дкЩњЮяФЄЬхЯЕжаЃЌЛљжЪЁЂЕчзгЪмЬхO2ЭЈЙ§ЩњЮяФЄБэУцЕФвКФЄБпНчВуЯђЩњЮяФЄФкВПРЉЩЂДЋЕнЁЃгЩгкРЉЩЂзшСІКЭЩњЮяЗДгІдьГЩЛљжЪХЈЖШЫцЩњЮяФЄЩюЖШБфЛЏЖјБфЛЏЃЌгыДЫЯргІЩњЮяФЄжаЮЂЩњЮяОњШКЫцЩюЖШГЪЗЧОљЯрЗжВМЁЃ
ЕБЩњЮяФЄдіМгЕНвЛЖЈКёЖШЪБЃЌдкФГвЛЩюЖШДІЃЌЕчзгЪмЬхЕФХЈЖШЮЊСуЁЃвдДЫЮЊНчЃЌЭтВуЕФЮЂЩњЮяПЩЭЌЪБЕУЕНЛљжЪКЭЕчзгЪмЬхЃЌНјаае§ГЃЕФЩњЮяЛЏбЇЗДгІЃЌЮЊЛюадВуЃЛФкВуЕФЮЂЩњЮядђгЩгкЕУВЛЕНЕчзгЪмЬхЃЌЦфКУбѕЩњЮяЛЏбЇЗДгІНЋжежЙЃЌЮЊЗЧЛюадВу(МћЭМ1)ЁЃ
|
|

вЛАуРДЫЕЃЌЛюадЩњЮяВуОЭЪЧОрЩњЮяФЄ—ЫЎНчУц100μmзѓгвЕФвЛВуЮЂЩњЮяЁЃЩњЮяФЄЕФдіГЄЪЕМЪЩЯЪЧЛюадВуЫцЪБМфдкДЙжБгкЩњЮяФЄ—ЫЎНчУцЗНЯђЩЯЯђЭтРЉеХЃЌЗЧЛюадВуж№НЅБфКёЕФЙ§ГЬЁЃдкЛюадВужаЃЌгЩгкВЛЭЌОњШКЖдЛљжЪЕФЧзКЭСІВЛЭЌвдМАдіжГЫйТЪЕФВювьЃЌЪЙЩњЮяФЄжаЕФзщЗждкИУВуЕФРЉеХЙ§ГЬжаВЛЖЯБфЛЏЁЃБОФЃаЭЕФжївЊФПЕФОЭЪЧУшЪідкЯѕЛЏЩњЮяФЄРЉеХЙ§ГЬжаЮЂЩњЮязщЗжЕФБфЛЏЙцТЩЁЃ
дкЩњЮяФЄЗДгІЦїжаЃЌЕБШмНтбѕХЈЖШдк2mg/LвдЯТЪБЃЌЯѕЛЏЗДгІЕФЫйТЪЪмШмНтбѕЫЎЦНЯожЦЁЃвРДЫЃЌИљОнжЪСПЪиКудРэЃЌЪзЯШНЈСЂЛюадВужаШмНтбѕЕФРЉЩЂ—ЗДгІЗНГЬЃЌЧѓНтГіЩњЮяФЄжаШмНтбѕЕФЗжВМЃЌШЛКѓИљОнШмНтбѕЗжВМЙцТЩЃЌЭЈЙ§ЩњЮяФЄЕФЩњГЄЗНГЬЭЦЕМГіЮЂЩњЮязщЗжвдМАЩњЮяФЄЕФРЉеХЫцЪБМфЕФБфЛЏЧщПіЁЃ
1.2ЛљБОМйЖЈ
дкФЃаЭЕФЭЦЕМЙ§ГЬжаМйЖЈЃК
ЂйЮЂЩњЮяПХСЃЮЊЧђаЮЃЌЩњЮяФЄдкЭЌвЛАыОЖЕФЧђУцЩЯОљдШЗжВМЁЃ
ЂкКіТдЭтВПДЋжЪЃЌМДВЛПМТЧвКФЄДЋжЪзшСІЁЃ
ЂлЛљжЪдкФЄФкДЋжЪЮЊЗжзгРЉЩЂЃЌКіТдЮЩСїЕШЦфЫћаЮЪНДЋжЪЁЃ
ЂмЩњЮяФЄЕФПеЯЖТЪЁЂУмЖШвдМАбѕЕФРЉЩЂЯЕЪ§ЃлЃИЃнЕШВЛЫцЩюЖШЗЂЩњБфЛЏЁЃ
ЂндкЮЂЩњЮяЕФдіжГЙ§ГЬжаВЛПМТЧЯИАћЕФФкдДДњаЛЃЌЮЂЩњЮяЖдЛљжЪЕФРћгУЗћКЯMonodФЃЪНЁЃ
ЂоВЛПМТЧЮЂЩњЮяФЄгЩгкЪмЕНСїЬхМєЧаКЭЩњЮяПХСЃжЎМфЕФФЅВСЖјЗЂЩњЕФЭбТфЁЃ
ЂпЩњЮяФЄжагЩгкЯѕЛЏЗДгІжЛЗЂЩњдкОрРыЩњЮяФЄБэУцКмНќЕФБЁВужаЃл8ЁЂ9ЃнЃЌвЛАуШЯЮЊИУВуКёЖШдМ50ЁЋ100μm,ЩѕжСгаЕФШЯЮЊНідк15ЁЋ20μmФкЯѕЛЏЮЂЩњЮяЕФЛюадНЯЧПЃл10ЃнЁЃвђДЫМйЖЈдкОрРыЩњЮяФЄ—ЫЎНчУцδОрРыФкЕФЩњЮяФЄгаЛюадЁЃ
ЂрдкМйЖЈЂпжаЕФδБЁВужаЃЌЩњЮяФЄЕФНсЙЙЁЂЮЂЩњЮяЕФГЩЗжЕШБЃГжВЛБфЃЌМДЛюадЩњЮяФЄЕФГЩЗжжЛЪЧЪБМфЕФКЏЪ§ЁЃ
ЂсЮТЖШБЃГжВЛБфЃЌМДИїжжЖЏСІбЇВЮЪ§ЮЊГЃЪ§ЁЃ
1.3ШмНтбѕРЉЩЂ—ЗДгІФЃаЭЕФНЈСЂ
гЩгкЛљжЪРЉЩЂЫљашвЊЕФЗДгІЪБМфБШЮЂЩњЮяЩњГЄЕФЬиеїЪБМфвЊаЁКУМИИіЪ§СПМЖЃЌЙЪЯрЖдгкЮЂЩњЮядіжГРДЫЕЛљжЪдкЩњЮяФЄжаЕФЗжВМПЩШЯЮЊДІгкЮШЖЈзДЬЌЁЃСэЭтЃЌРэТлКЭЪЕбщОљвбжЄУїЃЌЕБбѕАББШ<3.4ЪБЃЌШмНтбѕНЋГЩЮЊЗДгІЕФЯожЦадвђЫиЃл11ЃнЁЃвђДЫЃЌЕБвКЯржаШмНтбѕЕФХЈЖШ<2.5mg/LЪБЃЌбѕОЭЪЧЯѕЛЏЗДгІЕФЯожЦЛљжЪЃл1ЃнЁЃЫљвдЃЌдкЩњЮяФЄжаШмНтбѕЕФРЉЩЂ—ЗДгІЗНГЬЮЊЃК
|
ЁЁЁЁ |
ЪНжаDeff——ЛљжЪдкЩњЮяФЄжаЕФгааЇРЉЩЂЯЕЪ§ЃЌm2/s
Ci———ЛљжЪiЕФХЈЖШЃЌmg/L
r———ЩњЮяПХСЃАыОЖЃЌmm
ri——ЛљжЪiЕФЯћКФЫйТЪЃЌmg/(mgVSS.d)
бѕЕФБШЯћКФЫйТЪПЩвдШЯЮЊЪЧШмНтбѕХЈЖШЕФвЛМЖЗДгІЃЌМДЃК
|
ЁЁЁЁ |
ЪНжаqmax,NH4+,O2——АБбѕЛЏЗДгІжабѕЕФзюДѓБШЗДгІЫйТЪЃЌmgO2/ЃЈmgVSS.dЃЉ
qmax,NH2-,O2——бЧЯѕЫсбѕЛЏЗДгІжабѕЕФзюДѓБШЗДгІЫйТЪЃЌmgO2/(mgVSS.dЃЉ
Ko2,1——————АБЕЊЛЏЗДгІжабѕЦјЕФАыБЅКЭГЃЪ§
Ko2,2——————бЧЯѕЫсбѕЛЏЗДгІжабѕЦјЕФАыБЅКЭГЃ
pj———————ЩњЮяФЄжаЕкjжжЮЂЩњЮяЕФЬхЛ§жЪСП
fj———————ЩњЮяФЄжаЮЂЩњЮяОњШКjЫљеМЕФЬхЛ§ЗжТЪЃЌЖдгкЯѕЛЏЩњЮяФЄЯТБъ1жИАБЕЊЛЏОњЃЌЯТБъ2жИбЧЯѕЫсбѕЛЏОњ
ε———————ЩњЮяФЄЕФПеЯЖТЪ
ЪНжаpЁЂfjЖМжЛЪЧЪБМфЕФКЏЪ§ЃЌгыдкБЁВужаЕФЮЛжУЮоЙиЁЃИљОнМйЖЈЂсqmax,NO4+,O2ЁЂqmax,NO2-,O2ЁЂKo2,1КЭKo2,2ОљЮЊГЃЪ§ЁЃвђДЫЃЌЪНЃЈ2ЃЉПЩИФЮЊЃК
r02=KCO2(3)
ЪН(3)жаЃК
|
ЁЁ |
ЪНЃЈ1ЃЉПЩвдаДЮЊЃК
|
ЁЁЁЁ |
L——ЮЂЩњЮяФЄ——ЫЎНчУцДІЕФАыОЖЃЌmm
h——ЩњЮяФЄжаЮЂЩњЮяЕФжжРрЪ§
δ——ЛюадВуКёЖШЃЌmm
CO
2ЃЌp——вКЯржїЬхШмНтбѕХЈЖШЃЌmg/L
1.4ЩњЮяФЄдіжГРЉеХФЃаЭЕФНЈСЂ
ПМТЧдкtЪБПЬЛюадЩњЮяВужаЕкjжжЮЂЩњЮяЕФЬхЛ§ЗжТЪЮЊfjЃЌЩњЮяПХСЃЕФАыОЖЮЊLЃЌЖјдкt+dtЪБПЬЃЌЛюадЩњЮяВужаЕкjжжЮЂЩњЮяЕФЬхЛ§ЗжТЪЮЊfj+dfЃЌЩњЮяПХСЃЕФАыОЖБфЮЊL+dLЁЃдкdtЪБМфФкЛюадЩњЮяВужаЕкjжжЮЂЩњЮяЕФОЛдіжГЮЊЃК
 |
ЯТУцИљОнжЪСПЪиКуНЈСЂЩњЮяФЄРЉеХЗНГЬЃКдкdtЪБМфФкЛюадЩњЮяЕкjжжЮЂЩњЮяЕФжЪСПБфЛЏ=дкdtЪБМфФкЛюадЩњЮяВужаЕкjжжЮЂЩњЮяЕФОЛдіжГЃК
|
ЁЁЁЁ |
ЪНжаYj,o2——ЩњЮяФЄжаЕкjжжЮЂЩњЮявдбѕЦјБэЪОЕФЯИАћВњТЪЃЌmgVSS/mgO2
ЪНЃЈ6ЃЉМДЮЊЩњЮяФЄРЉеХЗНГЬЃЌНЋЦфМђЛЏЕУЃК
|
ЁЁЁЁ |
ЪНжаuL——БэУцЩњЮяФЄРЉеХЫйТЪЃЌm/sПЩвдБэЪОЮЊЃК
|
ЁЁЁЁ |
ЭЌЪБЃЌЩњЮяРАЛГвКЯрНчУцДІЕФАыОЖLПЩвдБэЪОЮЊЃК
|
ЁЁЁЁ |
ЪНжаro——дкЪБМфЮЊСуЪБЩњЮяПХСЃЕФАыОЖЃЌmm
ЖдгкЯѕЛЏЩњЮяФЄЃЌПМТЧЦфЩњЮяжжШКНіЮЊАБбѕЛЏОњКЭбЧЯѕбѕЛЏОњСНжжЃЌШчЙћФГвЛЪБПЬАБЕЊЛЏОњдкЮЂЩњЮяФЄжаЫљеМЕФЬхЛ§ЗжТЪf1=fЃЌдђбЧЯѕЫсбѕЛЏОњдкЮЂЩњЮяФЄжаЫљеМЕФЬхЛ§ЗжТЪf2=ЃЈ1-fЃЉЃЌНЋЪНЃЈ7ЃЉЁЂЃЈ8ЃЉЃЈ9ЃЉСЊСЂПЩвдаДГЩвдЯТаЮЪНЃК
|
ЁЁЁЁ |
СЊСЂЪН(5)ЁЂ(10)ЁЂ(11)ЃЌИљОнfКЭLГѕЪМжЕвдМАЯргІЕФВЮЪ§жЕ(Бэ1)ЃЌПЩвдЧѓГіЩњЮяФЄжаВЛЭЌжжРрЮЂЩњЮяЫљеМБШТЪКЭЩњЮяФЄРЉеХЫцЪБМфЕФБфЛЏЙиЯЕЁЃ
Бэ1МЦЫуЙ§ГЬжаВЮЪ§жЕ[2ЁЂ12]
|
2НсЙћгыЬжТл
2.1LгыfЫцЪБМфЕФБфЛЏ
ЭЈЙ§ЛљжЪНЕНтКЭЩњЮяФЄЩњГЄСНЙ§ГЬЕФМЦЫуЃЌПЩвдЕУГідквЛЖЈЬѕМўЯТЩњЮяФЄНсЙЙКЭЩњЮяПХСЃАыОЖЫцЪБМфЕФБфЛЏЙиЯЕЁЃЭМ2ИјГівКЯржїЬхШмНтбѕХЈЖШCp=0.5mg/LЃЌЛюадЩњЮяФЄКёЖШδ=50μmЪБЃЌАБбѕЛЏОњдкЩњЮяФЄжаЕФБШТЪfКЭЩњЮяПХСЃАыОЖLЫцЪБМфЕФБфЛЏЁЃ
ДгЭМ2ПЩвдПДГіЃЌАБбѕЛЏОњдкЩњЮяФЄжаЕФБШТЪЫцЪБМфЕФбгГЄж№НЅдіДѓЃЌвВОЭЪЧЫЕЫцзХЩњЮяФЄЕФРЉеХАБбѕЛЏОњНЋеМОнЯѕЛЏЩњЮяФЄБэУцЕФЮЛжУЃЌЪЙАБбѕЛЏЙ§ГЬЕФЗДгІЫйТЪдіДѓЃЛгЩгкбЧЯѕЫсбѕЛЏОњЕФБШТЪМѕЩйЃЌДгЖјЪЙбЧЯѕЫсбѕЛЏОњж№НЅЪЇШЅдкЩњЮяФЄжаЕФгаРћЮЛжУЃЌбЧЯѕЫсЕФбѕЛЏЫйТЪМѕаЁЃЌГіЫЎжаЕФNO2--NХЈЖШж№НЅдіМгЃЌжБжСЭъШЋбЧЯѕЫсЛЏЃЌетгыДѓЖрЪ§ЕФЪдбщЙлВьЭъШЋЗћКЯЁЃ
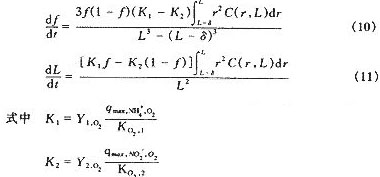 |
2.2CpЖдЛюадЩњЮяФЄжаОњШКЗжВМЕФгАЯь
гЩЪН(5)ЁЂ(10)КЭ(11)ПЩвдПДГіЃЌгАЯьЩњЮяФЄОњШКЗжВМБфЛЏЕФвђЫигаЛюадЩњЮяФЄЕФКёЖШδКЭвКЯржїЬхЕФбѕХЈЖШCpЁЃЭМ3ЮЊCpЖдАБбѕЛЏОњдкЛюадЩњЮяФЄжаЕФБШТЪfЕФгАЯьЁЃ
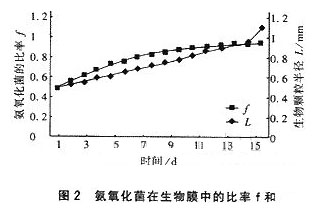 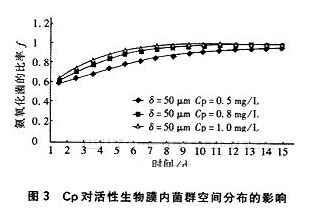 |
ДгЭМ3ПЩвдПДГіЃЌЕБЛюадЩњЮяФЄЕФКёЖШБЃГжВЛБфЪБЃЌвКЯржїЬхХЈЖШCpдНДѓдНгаРћгкАБбѕЛЏОњЕФОКељЩњГЄЃЌАБбѕЛЏОњдкНЯЖЬЕФЪБМфФкОЭПЩвдДяЕННЯИпЕФБШТЪЃЌвВОЭЪЧПЩвддкНЯЖЬЕФЪБМфФкЭъГЩАБбѕЛЏОњгыбЧЯѕЫсбѕЛЏОњжЎМфЕФбЁдёОКељЃЌДгЖјПЩдкНЯЖЬЕФЪБМфФкЪЕЯжГіЫЎбЧЯѕЫсбЮЕФИпЛ§РлЁЃвКЯржїЬхЕФШмНтбѕХЈЖШдНИпЃЌЭЈЙ§ЗжзгРЉЩЂНјШыЩњЮяФЄФкВПЬиЖЈЮЛжУЩЯЕФШмНтбѕХЈЖШвВОЭдНИпЃЌгЩгкАБбѕЛЏОњЖдШмНтбѕЕФЧзКЭСІБШбЧЯѕЫсбѕЛЏОњЖдШмНтбѕЕФЧзКЭСІвЊЧПЃЌЕМжТЕФАБбѕЛЏОњЩњГЄЫйТЪЕФдіМгБШбЧЯѕЫсбѕЛЏОњЩњГЄЫйТЪЕФдіМгвЊДѓЃЌЪЙОКељЙ§ГЬИќЮЊЯджјЁЃ
2.3δЖдЛюадЩњЮяФЄжаОњШКЗжВМЕФгАЯь
ЕБвКЯржїЬхХЈЖШЮЌГжвЛЖЈЪБЃЌЛюадЩњЮяФЄКёЖШδдНБЁЃЌдНШнвзЪЕЯжАБбѕЛЏОњКЭбЧЯѕЫсбѕЛЏОњЕФбЁдёОКељЃЌАБбѕЛЏОњОЭПЩвддкНЯЖЬЕФЪБМфФкеМОнЩњЮяФЄБэУцЕФЮЛжУЃЌЭъГЩЖдАБбѕЛЏОњЕФбЁдёЁЃетвВПЩвдгУАБбѕЛЏОњКЭбЧЯѕЫсбѕЛЏОњЖдШмНтбѕЕФЧзКЭСІВЛЭЌРДНтЪЭЃЌдквЛЖЈЕФШмНтбѕХЈЖШЬѕМўЯТЃЌЕБЛюадЩњЮяФЄНЯБЁЪБЃЌЦфжаЕФШмНтбѕЦНОљХЈЖШНЯДѓЃЌвВОЭШнвзЪЕЯжАБбѕЛЏОњЕФОКељЩњГЄЁЃ
3Нсгя
ЭЈЙ§НЈСЂЛљжЪРЉЩЂ—ЗДгІМАЮЂЩњЮяЩњГЄЕФЪ§бЇФЃаЭЃЌДгРэТлЩЯжЄЪЕСЫдкЯѕЛЏЩњЮяФЄжаДцдкзХАБбѕЛЏОњгыбЧЯѕЫсбѕЛЏОњжЎМфЖдПеМфКЭЛљжЪЕФОКељЁЃетжжОКељЪЧгЩгкЮЂЩњЮяБОЩэЕФдіжГЫйТЪВювьдьГЩЕФЁЃОКељЕФБиШЛНсЙћЪЧЫцзХЪБМфЕФбгајЃЌАБбѕЛЏОњдкЛюадЩњЮяФЄжаЕФБШТЪж№НЅдіДѓЃЌжБжСеМОнећИіЛюадВуЃЌДгЖјЪЕЯжГжОУЮШЖЈЕФбЧЯѕЫсЛЏЁЃ
ВЮПМЮФЯзЃК
Ѓл1ЃнGarridoJMetal.InfluenceofdissolvedoxygenconcentrationonnitriteaccumulationinabiofilmairliftsuspensionreactorЃлЃЪЃн.Biotech&Bioeng,1997,53:168-178.
Ѓл2ЃнPicioreanuCetal.ModelingtheeffectofoxygenconcentrationonnitriteaccumulationinabiofilmairliftsuspensionreactorЃлJЃн.WatSciTech,1997,36:147-156.
Ѓл3ЃнAblingU,SeyfriedCF.Anaerobic-aerobictreatmentofhighstrengthammoniumwastewaternitrogenremovalusingnitriteЃлJЃн.WatSciTech,1992,26:1007-1015.
Ѓл4ЃнЭѕЪРКЭЕШ.ЩњЮяСїЛЏДВНЕНтгаЛњЮяЕФЖЏСІбЇЛњРэбаОПЃлJЃн.ЖЋФЯДѓбЇбЇБЈ,1993ЃЌ23(1)ЃК62-68.Ѓл5ЃнFruhenMetal.SignificanceofspatialdistributionofmicrobialspeciesinmixedculturebiofilmsЃлJЃн.WatSciTech,1990,23:1365-1374.
Ѓл6ЃнReichertP,WannerO.Movementofsolidsinbiofilms:SignificanceofliquidphasetransportЃлJЃн.WatSciTech,1997,36:321-328.
Ѓл7ЃнWanner,ReichertP.Mathematicalmodelingofmixed-culturebiofilmsЃлJЃн.Biotech&Bioeng,1996,49:172-184.
Ѓл8ЃнPaulLBishop.BiofilmstructureandkineticsЃлJЃн.WatSciTech,1997,36:287-294.
Ѓл9ЃнScharmmAetal.StructureandfunctionofanitrifyingbiofilmasdeterminedbymicroelectrodesandfluorescentoligonucleotideprobesЃлJЃн.WatSciTech,1997,36:263-270.
Ѓл10ЃнLiuYuetal.SpecificactivityofnitrifyingbiofilminwaternitrificationprocessЃлJЃн.WatRes,1996,30:1645-1650.
Ѓл11ЃнShengKunetal.NitrificationanddenitrificationofhighstrengthammoniumandnitritewastewaterwithbiofilmreactorsЃлJЃн.WatSciTech,1991,23:1417-1425.
Ѓл12ЃнWiesmann.BiologicalnitrogenremovalfromwastewaterЃлAЃн.In:FiechterAdvancesinBiochemicalEngineering/BiotechnologhЃлЃУЃн,BerlinЃКSpringer-Verlag,1994,51.

ЪЙгУЮЂаХ“ЩЈвЛЩЈ”ЙІФмЬэМг“ЙШЬкЛЗБЃЭј”





